All animals have to deal with their world, and the objects in it. They walk on objects, crawl under them, avoid crashing into them, pick them up, eat them, mate with them, run away from them. Back in the geological dawn when evolution was young, animals had to make physical contact with objects before they could tell that those objects were there. What a bonanza of benefit was waiting for the first animal to develop a remote-sensing technology: awareness of an obstacle before hitting it; of a predator before being seized; of food that wasn't already within reach but could be anywhere in the large vicinity. What might this high technology be?
The sun provided not only the energy to drive the chemical cogwheels of life. It also offered the chance of a remote guidance technology. It pummelled every square millimeter of Earth's surface with a fusillade of photons: tiny particles travelling in straight lines at the greatest speed the universe allows, criss-crossing and ricocheting through holes and cracks so that no nook escaped, every cranny was sought out. Because photons travel in straight lines and so fast, because they are absorbed by some materials more than others and reflected by some materials more than others, and because they have always been so numerous and so all-pervading, photons provided the opportunity for remote-sensing technologies of enormous accuracy and power. It was necessary only to detect photons and - more difficult - distinguish the directions from which they came. Would the opportunity be taken up? Three billion years later you know the answer, for you can see these words.
Darwin famously used the eye to introduce his discussion on "Organs of extreme perfection and complication": To suppose that the eye, with all its inimitable contrivances for adjusting the focus to different distances, for admitting different amounts of light, and for the correction of spherical and chromatic aberration, could have been formed by natural selection, seems, I freely confess, absurd in the highest possible degree.
It is possible that Darwin was influenced by his wife Emma's difficulties with this very point. Fifteen years before The Origin of Species he had written a long essay outlining his theory of evolution by natural selection. He wanted Emma to publish it in the event of his death and he let her read it. Her marginalia survive and it is particularly interesting that she picked out his suggestion that the human eye may possibly have been acquired by gradual selection of slight but in each case useful deviations'. Emma's note here reads. 'A great assumption / E.D.' Long after The Origin of Species was published Darwin confessed, in a letter to an American colleague: 'The eye, to this day, gives me a cold shudder, but when I think of the fine known gradations, my reason tells me I ought to conquer the cold shudder.' Darwin's occasional doubts were presumably similar to those of the physicist whom I quoted at the beginning of Chapter 3. Darwin, however, saw his doubts as a challenge to go on thinking, not a welcome excuse to give up.
When we speak of the eye, by the way, we are not doing justice to the problem. It has been authoritatively estimated that eyes have evolved no fewer than forty times, and probably more than sixty times, independently in various parts of the animal kingdom. In some cases these eyes use radically different principles. Nine distinct principles have been recognized among the forty to sixty independently evolved eyes. I'll mention some of the nine basic eye types - which we can think of as nine distinct peaks in different parts of Mount Improbable's massif - as I go on.
How, by the way, do we ever know that something has evolved independently in two different groups of animals? For example, how do we know that bats and birds developed wings independently? Bats are unique among mammals in having true wings. It could theoretically be that ancestral mammals had wings, and all except bats have subsequently lost them. But for that to occur, an unrealistically large number of independent wing losses would be required, and the evidence supports common sense in indicating that this didn't happen. Ancestral mammals used their front limbs not for flying but for walking, as the majority of their descendants still do. It is by means of similar reasoning that people have worked out that eyes have arisen many times independently in the animal kingdom. We can also use other information such as details of how the eyes develop in the embryo. Frogs and squids, for instance. both have good camera-style eyes, but these eyes develop in such different ways in the two embryos that we can be sure they evolved independently. This does not mean that the common ancestor of frogs and squids totally lacked eyes of any kind. I wouldn't be surprised if the common ancestor of all surviving animals, who lived perhaps a billion years ago, possessed eyes. Perhaps it had some sort of rudimentary patch of light-sensitive pigment and could just tell the difference between night and day. But eyes, in the sense of sophisticated image-forming equipment, have evolved many times independently, sometimes converging on similar designs, sometimes coming up with radically different designs. Very recently there has been some exciting new evidence bearing upon this question of the independence of the evolution of eyes in different parts of the animal kingdom. I'll return to it at the end of the chapter.
As I survey the diversity of animal eyes, I'll often mention whereabouts on the slopes of Mount Improbable each type is to be found. Remember, though, that these are all eyes of modern animals, not of true ancestors. It is convenient to think that they might give us some clues about the kinds of eyes that ancestors had. They do at least show that eyes that we think of as lying half-way up Mount Improbable would actually have worked. This really matters for, as I have already remarked, no animal ever made a living by being an intermediate stage on some evolutionary pathway. What we may think of as a way station up the slope towards a more advanced eye may be, for the animal itself, its most vital organ and very probably the ideal eye for its own particular way of life. High resolution image-forming eyes, for instance, are not suitable for very small animals. High-quality eyes have to exceed a certain size - absolute size not size relative to the animal's body - and the larger the better in absolute terms. For a very small animal an absolutely large eye would probably be too costly to make and too heavy and bulky to carry around. A snail would look pretty silly if its eyes had the seeing power of human eyes (Figure 5.1). Snails that grew eyes even slightly larger than the present average might see better than their rivals. But they'd pay the penalty of having to carry a larger burden around, and therefore wouldn't survive so well. The largest eye ever recorded, by the way, is a colossal 37cm in diameter. The leviathan that can afford to carry such eyes around is a giant squid with 10-meter tentacles.
Accepting the limitations of the metaphor of Mount Improbable, let's go right down to the bottom of the vision slopes. Here we find eyes so simple that they scarcely deserve to be recognized as eyes at all. It is better to say that the general body surface is slightly sensitive to light. This is true of some single-celled organisms, some jellyfish, starfish, leeches and various other kinds of worms. Such animals are incapable of forming an image, or even of telling the direction from which light comes. All that they can sense (dimly) is the presence of (bright) light, somewhere in the vicinity. Weirdly, there is good evidence of cells that respond to light in the genitals of both male and female butterflies. These are not image-forming eyes but they can tell the difference be tween light and dark and they may represent the kind of starting point that we are talking about when we speak of the remote evolutionary origins of eyes. Nobody seems to know how the butterflies use them, not even William Eberhard, whose diverting book, Sexual Selection and Animal Genitalia, is my source for this information.
If we think of the plain below Mount Improbable as peopled by ancestral animals that were totally unaffected by light, the non directional light-sensitive skins of starfish and leeches (and butterfly genitals) are just a little way up the lower slopes, where the mountain path begins. It is not difficult to find the path. Indeed it may be that the plain of total insensitivity to light has always been small. It may be that living cells are more or less bound to be somewhat affected by light - a possibility that makes the butterflys light-sensitive genitals seem less strange. A light ray consists of a straight stream of photons. When a photon hits a molecule of some coloured substance it may be stopped in its tracks and the molecule changed into a different form of the same molecule. When this happens some energy is released. In green plants and green bacteria, this energy is used to build food molecules, in the set of processes called photosynthesis. In animals the energy may trigger a reaction in a nerve, and this constitutes the first step in the process called vision, even in animals lacking eyes that we would recognize as eyes. Any of a wide variety of coloured pigments will do, in a rudimentary way. Such pigments abound, for all sorts of purposes other than trapping light. The first faltering steps up the slopes of "Mount Improbable" would have consisted in the gradual improvement of pigment molecules. There is a shallow, continuous ramp of improvement - easy to climb in small steps.
This lowland ramp pushed on up towards the evolution of the living equivalent of the photocell, a cell specialized for capturing photons with a pigment, and translating their impact into nerve impulses. I shall continue to use the word photocell for those cells in the retina (in ourselves they are called rods and cones) which are specialized for capturing photons. The trick that they all use is to increase the number of layers of pigment available to capture photons. This is important because a photon is very likely to pass straight through any one layer of pigment and come out the other side unscathed. The more layers of pigment you have, the greater the chance of catching any one photon. Why should it matter how many photons are trapped and how many get through? Arent there always plenty of photons to spare? No, and the point is fundamental to our understanding of the design of eyes. There is a kind of economics of photons, an economics as mean-spirited as human monetary economics and involving inescapable trade-offs.
Before we even get into the interesting economic trade-offs, there can be no doubt that in absolute terms photons are in short supply at some times. On a crisp, starry night in 1986 I woke my two-year-old daughter juliet and carried her, wrapped in blankets, out into the garden where I pointed her sleepy face towards the published location of Halleys Comet. She didnt take in what I was saying, but I stubbornly whispered into her car the story of the comet and the certainty that I could never see it again but that she might when she was seventy~eight. I explained that I had woken her so that shed be able to tell her grandchildren in 2o62 that she had seen the comet before, and perhaps shed remember her father for his quixotic whim in carrying her out into the night to show it to her. (1 may even have whispered the words quixotic and whim because small children like the sound of words they dont know, carefully articulated.)
Probably some photons from Halleys Comet did indeed touchjuliets retinas that night in 1986 but, to be truthful, I had a hard time convincing myself that I could see the comet. Sometimes I seemed to conjure a faint, greyish smear at approximately the right place. At other times it melted away. The problem was that the number of photons falling on our retinas was close to zero. Photons arrive at random times, like raindrops. When it is really raining properly we are in no doubt of the fact and wish our umbrella hadnt been stolen. But when rain starts gradually, how do we decide the exact moment when it begins? We feel a single drop and look up apprehensively, unconvinced until a second or a third drop arrives. When rain is spitting infrequently like this, one person may say that it is raining while his companion denies it. The drops can fall infrequently enough to hit one person a minute before his companion registers a hit. To be really convinced that there is light, we need the photons to patter on our retinas at an appreciably high rate. When juliet and I gazed in the general direction of Halleys Comet, photons from the comet were probably hitting individual photocells on our retinas at the fantastically slow rate of about one every forty minutes! This means that any one photocell could be saying, Yes there is light there, while the vast majority of its neighbouring photocells were not. The only reason I received any sensation at all of a comet shaped object was that my brain was summing up the verdicts of hundreds of photocells. Two photocells capture more photons than one. Three capture more than two, and so on up the slope of "Mount Improbable". Advanced eyes like ours have millions of photocells densely packed like pile in a carpet, and each one of them is set up to capture as many photons as possible.
Figure 5.2 is a typical advanced photocell, from a human as it happens, but others are much the same. The writhing colony of apparent maggots in the middle of the picture are mitochondria. These are small bodies that live inside cells. They are originally descended from parasitic bacteria but they have made themselves indispensable for producing energy in all our cells. The nervous connecting wire of the photocell leads off at the left of the picture. The elegant rectangular array of membranes, lined up with military precision on the right, is where the photons are trapped. Each layer contains molecules of the vital, photon-trapping pigment. I count ninety-one layers of membrane in this picture. The exact number is not critical: the more the merrier as far as catching photons -is concerned, though there will be overhead costs prohibiting too many layers. The point is that ninety-one membranes are more effective in stopping photons than ninety, ninety are more effective than eighty nine, and so on back to one membrane, which is more effective than zero. This is the kind of thing I mean when I say that there is a smooth gradient up "Mount Improbable". We would be dealing with an abrupt precipice if, say, any number of membranes above forty-five was very effective while any number below forty-five was totally ineffective. Neither common sense nor the evidence leads us to suspect any such sudden discontinuity.
Squids, as we have seen, evolved their similar eyes independently of vertebrates. Even their photocells are very alike. The main difference is that in the squid the layers, instead of being packed as a stack of discs, are rings stacked around a hollow tube. This kind of superficial difference is common in evolution, for the same kind of inconsequential reason as, say, the fact that English light switches go down for on, American ones down for off. All advanced animal photocells are playing different versions of the same trick of increasing the number of layers of pigment laced membranes through which a photon must pass if it would escape untrapped. From Mount Improbables point of view, the important point is that one more layer marginally improves the chance of trapping photons no matter how many, or how few, layers there already are. Ultimately, when most of the photons have been caught, there will be a law of diminishing returns for the increasing cost of more layers.
Of course there is not much call, in the wild, for detecting Halleys Comet, returning every seventy-six years with its negligible contribution of reflected photons. But it is very useful to have eyes sensitive enough to see by moonlight and even starlight if you are an owl. On a typical night, any one of our photocells might receive photons at about one per second, admittedly a higher rate than for the comet, but still slow enough to make it vital to trap every last photon if it can be done. But when we speak of a harsh economics of photons it would be quite wrong to assume that the harshness is confined to the night. In bright sunlight the photons may drum the retina like a tropical cloudburst, but there is still a problem. The essence of seeing a patterned image is that photocells in different parts of the retina must report different intensities of light and this means distinguishing different rates of pattering in different parts of the photon rainstorm. The sorting of photons coming from different fine-grained parts of the scene can lead to local impoverishments of photons just as serious as the global impoverishment of the night. It is to this sorting that we now turn.
Photocells on their own just tell an animal whether there is light or not. The animal can tell the difference between night and day, and can tell when a shadow falls which might, for example, portend a predator. The next step of improvement must have been the acquisition of some rudimentary sensitivity to direction of light and direction of movement, of say, a menacing shadow. The minimal way of achieving this is to back the photocells with a dark screen on one side only. A transparent photocell without a dark screen receives light from all directions and cannot tell where light is coming from. An animal with only one photocell in its head can steer towards, or away from, light, provided the photocell is backed by a screen. A simple recipe for doing this is to swing the head like a pendulum from side to side; if the light intensity on the two sides is unbalanced, change direction until it is balanced. There are some maggots that follow this recipe for steering directly away from light.
But swinging your head from side to side is a rudimentary way of detecting the direction of light, fit for the lowest slopes of "Mount Improbable". A better way is to have more than one photocell pointing in different directions, each one backed by a dark screen. Then by comparing the rates of photon rain on the two cells you can make inferences about direction of light. If you have a whole carpet of photocells, a better way is to bend the carpet, with its backing screen, into a curve, so that the photocells on different parts of the curve are pointing in systematically different directions. A convex curve can give rise, eventually, to the sort of compound eye that insects have, and Ill return to this. A concave curve is a cup and it gives rise to the other main kind of eye, the camera eye like our own. Photocells in different parts of a cup will fire when light is coming from different directions, and the more cells there are the finer-grained will be the discrimination.
The light rays (parallel white lines with arrows) are halted by the thick black screen at the back of the cup (Figure 5.3). By keeping track of which photocells are firing and which are not, the brain can detect the direction from which the light is coming. From the point of view of climbing "Mount Improbable", what matters is that there is a continuous evolutionary gradation, a smooth incline up the mountain, connecting animals with a flat carpet of photocells to animals with a cup. Cups can get gradually deeper or gradually shallower, by continuous slow degrees. The deeper the cup, the greater the ability of the eye to discriminate light coming from different directions. On the mountain, no steep precipices have to be leapt.
Cup eyes like this are common in the animal kingdom. Figure 5.4 shows the eye of a limpet, of a bristleworm, of a clam and of a flatworm. These eyes have probably evolved their cup shape independently. This is particularly clear in the case of the flatworm eye which betrays its separate origin by keeping its photocells inside the cup. On the face of it this is an odd arrangement - the light rays have to penetrate a thicket of connecting nerves before they hit the photocells - but lets not be snobbish about it because the same apparently poor design mars our own much more sophisticated eyes. Ill return to this and show that it isnt really such a bad idea as it seems.
In any case, a cup eye on its own is far from capable of forming what we humans, with our excellent eyes, would recognize as a proper image. Our kind of image-formation, which depends upon the lens principle, needs a little explanation. We approach the problem by asking why an unaided carpet of photocells, or a shallow cup, will not see an image of, say, a dolphin, even when the dolphin is conspicuously displayed in front of it.
If light rays behaved as in Figure 5.5, everything would be easy and an image of the dolphin, the right way up, would appear on the retina. Unfortunately they dont. To be more precise, there are rays that do exactly what I have drawn in the picture. The trouble is that these are swamped by any number of rays going in every other direction at the same time. Every bit of the dolphin sends a ray to every point on the retina. And not just every bit of the dolphin, but every bit of the background and of everything else in the scene. You can think of the result as an infinite number of dolphin images, in every possible position on the surface of the cup and every possible way up and way round. But what this amounts to, of course, is no image at all, just a smooth spreading of light over the whole surface (Figure 5.6).
We have diagnosed the problem. The eye is seeing too much: an infinity of dolphins instead of only one. The obvious solution is to subtract: cut out every dolphin image except one. It wouldnt matter which one, but how to get rid of the rest? One way is to trudge on up the same slope of "Mount Improbable" as gave us the cup, steadily deepening and enclosing the cup until the aperture has narrowed to a pinhole. Now the vast majority of rays are prevented from entering the cup. The minority that remain are just those rays that form a small number of similar images - upside-down - of the dolphin ( Figure 5.7). If the pinhole becomes extremely small, the blurring disappears and a single, sharp picture of the dolphin remains (actually, extremely small pinholes introduce a new kind of blurring, but well forget about that for a moment). You can think of the pinhole as an image filter, removing all but one of the bewildering visual cacophony of dolphins.
The pinhole effect is just an extreme version of the cup effect that we have already met as an aid to telling the direction of light. It belongs only a bit farther up the same slope of "Mount Improbable" and there are no sharp precipices between. There is no difficulty in a pinhole eyes evolving from a cup eye, and no difficulty in a cup eyes evolving from a flat sheet of photocells. The slope up the mountain from flat carpet to pinhole is gradual and easily climbable all the way. Climbing it represents a progressive knocking out of conflicting images until, at the peak, only one is left.
Pinhole eyes in varying degrees are, indeed, scattered around the animal kingdom. The most thoroughgoing pinhole eye is that of the enigmatic mollusc Nautilus (Figure 5.8a), related to the extinct ammonites (and a more distant relative of an octopus, but with a coiled shell). Others, such as the eye of a marine snail in Figure 5.8b, are perhaps better described as deep cups than true pinholes. They illustrate the smoothness of this particular gradient climbing up "Mount Improbable".
A first thought suggests that the pinhole eye ought to work rather well, provided you make the pinhole small enough. If you make the pinhole almost infinitely small, you might think that youd get an almost infinitely perfect image by cutting out the vast majority of competing, interfering images. But now two new snags arise. One is diffraction. I deferred talking about it just now. It is a blurring problem that results from the fact that light behaves like waves, which can interfere with each other. This blurring gets worse when the pinhole is very small. The other snag with a small pinhole recalls the hard trade-offs of our photon economy. When the pinhole is small enough to make a sharp image, it necessarily follows that so little light gets through the hole that you can see the object well only if it is illuminated by an almost unattainably bright light. At normal lighting levels not enough photons get through the pinhole for the eye to be certain what it is seeing. With a tiny pinhole we have a version of the Halleys Comet problem. You can combat this by opening out the pinhole again. But now you are back where you were with a confusion of competing dolphins. The photon economy has brought us to an impasse on this particular foothill of "Mount Improbable". With the pinhole design you can have a sharpish but dark image, or a brightish but fuzzy one. You cannot have both. Such trade-offs are meat and drink to economists, which is why I coined the notion of an economy of photons. But is there no way to achieve a bright and yet simultaneously sharp image? Fortunately there is.
First, think of the problem computationally. Imagine that we broaden the pinhole out to let in a nice lot of light. But instead of leaving it as a gaping hole, we insert a magic window, a masterpiece of electronic wizardry embedded in glass and connected to a computer (Figure 5.9). The property of this computer-controlled window is the following. Light rays, instead of passing straight through the glass, are bent through a cunning angle. This angle is carefully calculated by the computer so that all rays originating from a point (say the dolphins nose) are bent to converge on a corresponding point on the retina. Ive drawn only the rays from the dolphins nose, but the magic screen, of course, has no reason to favor any one point and does the same calculation for every other point as well. All rays originating at the tail of the dolphin are bent in such a way that they converge on a corresponding tail point on the retina, and so on. The result of the magic window is that a perfect image of the dolphin appears on the retina. But it is not dark like the image from a tiny pinhole, because lots of rays (which means torrents of photons) converge from the nose of the dolphin, lots of rays converge from the tail of the dolphin, lots of rays converge from every point on the dolphin to their own particular point on the retina. The magic window has the advantages of a pinhole, without its great disadvantage.
Its all very well to conjure up a so-called magic window out of imaginative thin air. But isnt it easier said than done? Think what a complicated calculation the computer attached to the magic window is doing. It is accepting millions of light rays, coming from millions of points out in the world. Every point on the dolphin is sending millions of rays at millions of angles to different points on the surface of the magic window. The rays are criss-crossing one another in a bewildering spaghetti junction of straight lines. The magic window with its associated computer has to deal with each of these millions of rays in turn and calculate its own particular angle, through which it must be precisely turned. Where is this wonderful computer to come from, if not from a complicated miracle? Is this where we meet our Waterloo: an inevitable precipice in our ascent up "Mount Improbable"?
Remarkably, the answer is no. The computer in the diagram is just an imaginary creation to emphasize the apparent complexity of the task if you look at it in one way. But if you approach the problem in another way the solution turns out to be ludicrously easy. There is a device of preposterous simplicity which happens to have exactly the properties of our magic window, but with no computer, no electronic wizardry, no complication at all. That device is the lens. You dont need a computer because the calculations need never be done explicitly at all. The apparently complicated calculations of millions of ray angles are taken care of, automatically and without fuss, by a curved blob of transparent material. Ill take a little time to explain how lenses work, as a prelude to showing that the evolution of the lens wouldnt have been very difficult.
It is a fact of physics that light rays are bent when they pass from one transparent material into another transparent material (Figure 5.10). The angle of bending depends upon which two materials they happen to be, because some substances have a greater refractive index - a measure of the capacity to bend light - than others. If we are talking about glass and water, the angle of bending is slight because the refractive index of water is nearly the same as that of glass. If the junction is between glass and air, the light is bent through a bigger angle because air has a relatively low refractive index. At the junction between water and air, the angle of bending is substantial enough to make an oar look bent.
Figure 5.10 represents a block of glass in air. The thick line is a light ray entering the block, being bent within the glass, then bending back to the original angle as it goes out the other side. But of course there is no reason why a blob of transparent material should have neatly parallel sides. Depending upon the angle of the surface of the blob, a ray can be sent off in any direction you choose. And if the blob is covered with facets at lots of different angles, a set of rays can be sent off in lots of different directions (Figure 5.11). If the blob is curved convexly on one or both of its sides, it will be a lens: the working equivalent of our magic window. Transparent materials are not particularly rare in nature. Air and water, two of the commonest substances on our planet, are both transparent. So are many other liquids. So are some crystals if their surface is polished, for instance by wave action in the sea, to remove surface roughness. Imagine a pebble of some crystalline material, worn into a random shape by the waves. Light rays from a single source are bent in all sorts of directions by the pebble, depending upon the angles of the pebbles surfaces. Pebbles come in all sorts of shapes. Quite commonly they are convex on both sides. What will this do to light rays from a particular source like a light bulb?
When the rays emerge from a pebble with vaguely convex sides they will tend to converge. Not to a neat, single point such as would reconstruct a perfect image of the light source like our hypothetical magic window. That would be too much to hope. But there is a definite tendency in the right direction. Any quartz pebble whose weathering happened to make it smoothly curvaceous on both sides would serve as a good magic window, a true lens capable of forming images which, though far from sharp, are much brighter than a pinhole could produce. Pebbles worn by water usually are, as a matter of fact, convex on both sides. If they happened to be made of transparent material many of them would constitute quite serviceable, though crude, lenses.
A pebble is just one example of an accidental, undesigned object which can happen to work as a crude lens. There are others. A drop of water hanging from a leaf has curved edges. It cant help it. Automatically, without further design from us, it will function as a rudimentary lens. Liquids and gels fall automatically into curved shapes unless there is some force, such as gravity, positively opposing this. This will often mean that they cannot help functioning as lenses. The same is often true of biological materials. A young jellyfish is both lens-shaped and beautifully transparent. It works as a tolerably good lens, even though its lens properties are never actually used in life and there is no suggestion that natural selection has favored its lens-like properties. The transparency probably is an advantage because it makes it hard for enemies to see, and the curved shape is an advantage for some structural reason having nothing to do with lenses.
Here are some images I projected on to a screen using various crude and undesigned image- forming devices. Figure 5.12a shows a large letter A, as projected on a sheet of paper at the back of a pinhole camera (a closed cardboard box with a hole in one side). You probably could scarcely read it if you werent told what to expect, even though I used a very bright light to make the image. In order to get enough light to read it at all, I had to make the pin hole quite large, about a centimeter across. I might have sharpened the image by narrowing the pinhole, but then the film would not have registered it the familiar trade-off we have already discussed.
Now see what a difference even a crude and undesigned lens makes. For Figure 5.12b the same letter A was again projected through the same hole on to the back wall of the same cardboard box. But this time I hung a polythene bag filled with water in front of the hole. The bag was not designed to be particularly lens-shaped. It just naturally hangs in a curvaceous shape when you fill it with water. I suspect that a jellyfish, being smoothly curved instead of bucked up into creases, would have produced an even better image. Figure 5.12c (CAN YOU READ THIS?) was made with the same cardboard box and hole, but this time a round wine goblet filled with water was placed in front of the hole instead of a sagging bag. Admittedly the wineglass is a man-made object, but its designers never intended it to be a lens and they gave it its globular shape for other reasons. Once again, an object that was not designed for the purpose turns out to be an adequate lens.
Of course polythene bags and wineglasses were not available to ancestral animals. I am not suggesting that the evolution of the eye went I -, through a polythene-bag stage, any more than it went through a cardboard-box stage. The point about the polythene bag is that, like a raindrop or a jellyfish or a rounded quartz crystal, it was not designed as a lens. It takes on a lens-like shape for some other reason which happens to be influential in nature.
It is not difficult, then, for rudimentary lens-like objects to come into existence spontaneously. Any old lump of half-way transparent jelly need only assume a curved shape (there are all sorts of reasons why it might) and it will immediately confer at least a slight improvement over a simple cup or pinhole. Slight improvement is all that is required to inch up the lower slopes of "Mount Improbable". What might the intermediates have looked like? Look back at Figure 5.8, and once again I must stress that these animals are modern and must not be thought of as an actual ancestral series. Notice that the cup in Figure 5.8b (marine snail) has a lining of transparent jelly, the vitreous mass (vm) which perhaps serves to protect the sensitive photocells from the raw sea water which flow: freely through the aperture into the cup. That purely protective vitreous mass has one of the necessary qualities of a lens - transparency - but it lacks the correct curvature and it needs thickening up. Now look at Figures 5.8c, 5.8d and 5.8e, eyes from a bivalve mollusk, an abalone and a ragworm. In addition to providing yet more examples of cups and intermediates between cups and pinholes, all these eyes show greatly thickened vitreous masses. Vitreous masses, of varying degrees of shapelessness, are ubiquitous in the animal kingdom. As a lens, none of those splodges of jelly would move Mr Zeiss or Mr Nikon to write home. Nevertheless, any lump of jelly that has a little convex curvature would mark significant improvements over an open pinhole.
The biggest difference between a good lens and something like the abalones vitreous mass is this: for best results the lens should be detached from the retina and separated from it by some distance.. The gap need not be empty. It could be filled by more vitreous mass. What is needed is that the lens should have a higher refractive index than the substance that separates the lens from the retina. There are various ways in which this might be achieved, none of them difficult. I'll deal with just one way, in which the lens is condensed from a local region within the front portion of a vitreous mass like that in Figure 5.8e.
First, remember that a refractive index is something that every transparent substance has. It is a measure of its power to bend rays of light. Human lens-makers normally assume that the refractive index of a lump of glass is uniform through the glass. Once a ray of light has entered a particular glass lens and changed direction appropriately, it goes in a straight line until it hits the other side of the lens. The lens-makers art lies in grinding and polishing the surface of the glass into precision shapes, and in joining different lenses together in compound cascades.
You can glue different kinds of glass together in complicated ways to make compound lenses with lots of different refractive indexes in various parts of them. The lens in Figure 5.13a, for instance, has a central core made of a different kind of glass with a higher refractive index. But there are still discrete changes from one refractive index to another. In principle, however, there is no reason why a lens should not have a continuously varying refractive index throughout its interior. This is illustrated in Figure 5.13b. This graded index lens is hard for human lens-makers to achieve because of the way they make their lenses out of glass. After writing this I was informed by a correspondent, Howard Kleyn, formerly of the Cable and Wireless Company, that humans do, as a matter of fact, make something equivalent to a graded index lens. It is actually a graded index optical fibre. By his description, it works like this. You start with a hollow tube of good glass, about a meter long and a few centimeters in diameter, which you heat up. You then puff into the tube finely powdered glass. The powdered glass melts and fuses with the lining of the tube, thereby thickening the lining while narrowing the bore of the tube. Now conics the cunning part. As this procedure progresses, the powder that is puffed in is of gradually changing quality: specifically, it has been ground from glass of progressively increasing refractive index. By the time the hollow bore has narrowed to nothing, the tube has become a rod made of highly refracting glass at its central core with gradually decreasing refractive index as you move towards its outer layers. The rod is then heated again, and drawn out into a fine filament. This filament retains the same graded refractive index, from core to periphery, in miniature, as the rod from which it was drawn. It is technically a graded index lens, albeit a very thin, long one. its lens property is used not for focusing an image but for improving its quality as a light guide which does not allow its beam of light to disperse. Several of these filaments would normally be used to manufacture a multi-stranded optical fibre cable. But it is easy for living lenses to be built like this because they are not made all at one time: they grow from small beginnings as the young animal develops. And, as a matter of fact, lenses with continuously varying refractive indexes are found in fish, octopuses and many other animals. If you look carefully at Figure 5.8e, you see what might conceivably be a region of differing refractive index in the zone behind the aperture of the eye.
But I was starting to tell the story of how lenses might have evolved in the first place, from a vitreous mass that filled the whole eye. The principle of how it might have happened, and the speed with which it might have been accomplished, has been beautifully demonstrated in a computer model by a pair of Swedish biologists called Dan Nilsson and Susanne Pelger. I shall lead up to explaining their elegant computer model in a slightly oblique way. Instead of going straight to what they actually did, I shall return to our progression from Biomorph to NetSpinner computer models and ask how one could ideally set about making a similar computer model of the evolution of an eye. I shall then explain that this is essentially equivalent to what Nilsson and Pelger did, although they didnt put it in quite the same way.
Recall that the biomorphs evolved by artificial selection: the selecting agent was human taste. We couldnt think of a realistic way of incorporating natural selection into the model so we switched to model spider webs instead. The advantage of spider webs was that, since they do their work in a two-dimensional plane, their efficiency in catching flies could be calculated by the computer automatically. So could their cost in silk, and model webs could therefore be automatically chosen by the computer in a form of natural selection. We agreed that spider webs were exceptional in this respect: we could not easily hope to do the same for the backbone of a hunting cheetah or the fluke of a swimming whale, because the physical details involved in assessing a three dimensional organs efficiency are too complicated. But an eye is like a spider web in this respect. The efficiency of a model eye drawn in two dimensions can be assessed automatically by the computer. I am not implying that an eye is a two-dimensional structure, because it isnt. It is just that, if you assume that the eye is circular when seen head on, its efficiency in three dimensions can be assessed from a computer picture of a single vertical slice through the middle. The computer can do a simple ray-tracing analysis and work out the sharpness of image that an eye would be capable of forming. This quality scoring is equivalent to NetSpinners calculation of the efficiency of a computer spider web at catching computer flies.
Just as NetSpinner webs procreated mutant daughter webs, so we could let model eyes generate mutant daughter eyes. Each daughter eye would have basically the same shape as the parent, but with a small random change to some minor aspect of its shape. Of course some of these computer eyes would be so unlike real eyes as not to deserve the title, but no matter. They could still be bred, and their optical quality could still be given a numerical score - presumably it would be very low. We could therefore, in the same way as NetSpinner, evolve improved eyes by natural selection in the computer. We could either start with a fairly good eye and evolve a very good eye. Or we could start with a very poor eye or even with no eye at all.
It is instructive to run a program like NetSpinner as an actual simulation of evolution, setting it off from a rudimentary starting point and waiting to see where it will end up. You could even end up at different culmination points on different evolutionary runs, because there could be alternative accessible peaks of "Mount Improbable". We could run our eye model in evolution mode too, and it would make a vivid demonstration. But actually you dont learn much more by letting the model evolve than you would learn by exploring, more systematically, where the upward path(s) on "Mount Improbable" lead(s). From a given starting point, a path which goes ever upward, never downward, is the path that natural selection would follow. If you ran the model in evolutionary mode, natural selection would follow that path. So it saves computer time if we search systematically for upward paths and for peaks that can be reached from.postulated starting points. The important thing is that the rules of the game forbid going downhill. This more systematic search for upward paths is what Nilsson and Pelger did, but you can see why I chose to introduce their work as if we were planning, with them, a NetSpinner-style enactment of evolution.
However we choose to run our model, whether in natural-selection mode or in systematic exploration of the mountain mode, we have to decide upon some rules of embryology: that is, some rules governing how genes control the development of bodies. What aspects of shape do the mutations actually operate upon? And how big, or how small, are the mutations themselves? In the case of NetSpinner, the mutations act upon known aspects of the behavior of spiders. In the case of biomorphs, mutations act upon the lengths and angles of branches in growing trees. In the case of eyes, Nilsson and Pelger began by acknowledging that there are three main types of tissue in a typical camera eye. There is an outer casing to the camera, usually opaque to light. There is a layer of light-sensitive photocells. And there is some kind of transparent material, which may serve as a protective window or which may fill the cavity inside the cup, if indeed there is a cup, for we are not taking anything for granted in our simulation. Nilsson and Pelgers starting point - the foot of the mountain - is a flat layer of photocells (grey in Figure 5.14), sitting on a flat backing screen (black) and topped by a flat layer of transparent tissue (off-white). They assumed that mutation works by causing a small percentage change in the size of something, for example a small percentage decrease in the thickness of the transparent layer, or a small percentage increase in the refractive index of a local region of the transparent layer. Their question really is, where can you get to on the mountain if you start from a given base camp and go steadily upwards? Going upwards means mutating, one small step at a time, and only accepting mutations that improve optical performance.
So, where do we get to? Pleasingly, through a smooth upward pathway, starting from no proper eye at all, we reach a familiar fish eye, complete with lens. The lens is not uniform like an ordinary man-made lens. It is a graded index lens such as we met in Figure 5.13b. Its continuously varying refractive index is represented in the diagram by varying shades of grey. The lens has condensed out of the vitreous mass by gradual, point by point changes in the refractive index. There is no sleight of hand here. Nilsson and Pelger didnt pre-program their simulated vitreous mass with a primordial lens just waiting to burst forth. They simply allowed the refractive index of each small bit of transparent material to vary under genetic control. Every smidgen of transparent material was free to vary its refractive index in any direction at random. An infinite number of patterns of varying refractive index could have emerged within the vitreous mass. What made the lens come out lensshaped was unbroken upward mobility, the equivalent of selectively breeding from the best seeing eye in each generation.
Nilsson and Pelgers purpose was not only to show that there is a smooth trajectory of improvement from a flat non-eye to a good fish eye. They were also able to use their model to estimate the time it would take to evolve an eye from nothing. The total number of steps that their model took was 1 829 if each step achieved a i% change in the magnitude of something. But there is nothing magic about 1%. The same total quantity of change would have taken 363 992 steps of 0.01%. Nilsson and Pelger had to re-express the total quantity of change in non-arbitrary, realistic units, and that means units of genetic change. In order to do this, it was necessary to make some assumptions. For example they had to make an assumption about the intensity of selection. They assumed that for every 101 animals possessing an improved eye who survived, 100 animals without the improvement survived. As you can see, this is a low intensity of selection as common sense might judge it - you are almost as well off without the improvement as with it. They deliberately chose a low, conservative or pessimistic figure because they were bending over backwards to bias their estimate of rate of evolution towards being, if anything, too slow. They also had to make two other assumptions: of heritability and of coefficient of variation. The coefficient of variation is a measure of how much variation there is in the population. Natural selection needs variation to work upon and Nilsson and Pelger again deliberately chose a pessimistically low value. Heritability is a measure of how much of the variation, out of a given populations available variation, is inherited. If the heritability is low it means that most of the variation in the population is environmentally caused, and natural selection, for all that it may choose individuals to live or die, will have little impact on evolution. If heritability is high, selection.,has a large impact on future generations because individual survival really translates into gene survival. Heritabilities frequently turn out to be more than 50%, so Nilsson and Pelgers decision to settle on 50% was a pessimistic assumption. Finally, they made the pessimistic assumption that different parts of the eye could not change simultaneously in one generation.
Pessimistic in all these cases means that the estimate that we finally come up with for how long it takes an eye to evolve is likely to be on the long side. The reason we call an over-estimate pessimistic rather than optimistic is this. A sceptic about the power of evolution, such as Emina Darwin, is naturally drawn to the view that an organ as notoriously complicated and many-parted as an eye, if it can evolve at all, will take an immense time to evolve. Nilsson and Pelgers final estimate was actually astoundingly short. At the end of the calculation, it turned out that it would take only about 364 000 generations to evolve a good fish eye with a lens. It would have been even shorter if they had made more optimistic (and this probably means more realistic) assumptions.
How long is 364 000 generations in years? That depends on the generation time, of course. The animals we are talking about would he small marine animals like worms, mollusks and small fish. For them, a generation typically takes one year or less. So Nilsson and Pelgers conclusion is that the evolution of the lens eye could have been accomplished in, less than half a million years. And that is a very very short time indeed by geological standards. It is so short that, in the strata of the ancient eras we are talking about, it would be indistinguishable from instantaneous. The plaint that there hasn't been enough time for the eye to evolve turns out to be not just wrong but dramatically, decisively, ignominiously wrong.
Of course there are some other details of a full-fledged eye that Nilsson and Pelger have not yet dealt with and which might (though they dont think so) take rather longer to evolve. There is the preliminary evolution of the light-sensitive cells - what I have been calling photocells - which they regarded as having been accomplished before the start of their model evolution system. There are other, advanced features of modem eyes such as the apparatuses for changing the focus of an eye, for changing the size of the pupil or f-stop, and for moving the eye. There are also all the systems in the brain that are needed for processing the information from the eye. Moving the eye is important, not just for the obvious reason but, more indispensably, to hold the gaze still while the body moves. Birds do this by using the neck muscles to keep the whole head still, notwithstanding substantial movements of the rest of the body. Advanced systems for doing this involve quite sophisticated brain mechanisms. But it is easy to see that rudimentary, imperfect adjustments would be better than nothing, so there is no difficulty in piecing together an ancestral series following a smooth path up "Mount Improbable".
In order to focus rays that are coming from a very distant target, you need a weaker lens than to focus rays that are coming from a close target. To focus sharply both far and near is a luxury one can live without, but in nature every little boost to the chances of survival counts and as a matter of fact different sorts of animals display a variety of mechanisms for changing the focus of the lens. We mammals do it by means of muscles that pull on the lens and change its shape a little. So do birds and most reptiles. Chameleons, snakes, fishes and frogs do it in the same way a camera does, by pulling the lens a little way forwards or backwards. Animals with smaller eyes dont bother. Their eyes are like a Box Brownie: approximately, though not brilliantly, in focus at all distances. As we get older our eyes sadly become more Box Brownie-like and we often need bifocal glasses to see both near and far.
It is not at all difficult to imagine the gradual evolution of mechanisms for changing focus. When experimenting with the polythene bag filled with water, I quickly noticed that the sharpness of focus could be made better (or worse) by poking the bag with my fingers. Without being consciously aware of the shape of the bag, without even looking at the bag but concentrating on the quality of the image being projected, I simply poked and squashed the bag at random until the focus got better. Any muscle in the vicinity of a lump of vitreous mass could, as a byproduct of contracting for some other purpose, incidentally improve the focus of the lens. This opens up a broad highway for gentle improvement all the way up the slopes of "Mount Improbable", which could culminate in either the mammal or the chameleon method of changing the focus.
Changing the aperture - the size of the hole through which light is admitted - may be slightly more difficult, but not much. The reason for wanting to do this is the same as in a camera. For any given sensitivity of film/photocells, it is possible to have too much light (dazzle) as well as too little. Moreover, the narrower the hole, the better the depth of focus - the range of distances that are simultaneously in focus. A sophisticated camera, or eye, has a built-in light meter which automatically stops down the hole when the sun comes out, and opens up the hole when the sun goes in. The pupil of a human eye is a pretty sophisticated piece of automation technology, something that Japanese micro-engineer could be proud of.
But, once again, it isn't difficult to see how this advanced mechanism might have got its start on the lower slopes of "Mount Improbable". We think of the pupil as circular, but it doesnt have to be. Any shape would do (see different pupils). Sheep and cattle have a long, horizontal, lozenge-shaped pupil. So do octopuses and some snakes, but other snakes have a vertical slit. Cats have a pupil which varies from a circle to a narrow, vertical slit (Figure 5.15c):
Does Minnaloushe know that his pupils will pass from change to change,
and that from round to crescent, from crescent to round they range?
Minnaloushe creeps through the grass alone, important and wise,
and lifts to the changing moon his changing eyes.
W. B. Yeats
Even expensive cameras often have pupils which are crude polygons rather than perfect circles. All that matters is that the quantity of light entering the eye should be controlled. When you realize this, the early evolution of the variable pupil ceases to be a problem. There are lots of gentle paths to be followed up the lower slopes of "Mount Improbable". The iris diaphragm is no more an impenetrable evolutionary barrier than is the anal sphincter. Perhaps the most important quantity that needs to be improved is the speed of responsiveness of the pupil. Once you have nerves at all, speeding them up is an easy glide up the slopes of the mountain. Human pupils respond fast, as you can quickly verify by shining a torch in your eye while looking at your pupil in a mirror. (you see the effect most dramatically if you shine the torch in one eye while looking at the pupil in the other: for the two are ganged together.)
As weve seen, the Nilsson and Pelger model developed a graded index lens, which is different from most man-made lenses but like those of fishes, squids and other underwater camera eyes. The lens arises by condensation of a zone of locally high-refractive index within previously uniform transparent jelly.
Not all lenses evolved by condensing out from a gelatinous mass. Figure 5.16 shows two insect eyes that form their lenses in quite different ways. These are both so-called simple eyes, not to be confused with the compound eyes which well come to in a moment. In the first of these simple eyes, from a sawfly larva, the lens forms as a thickening of the cornea - the outer transparent layer. In the second one, from a mayfly, the cornea is not thickened and the lens develops as a mass of colorless, transparent cells. Both these two methods of lens development lend themselves to the same kind of "Mount Improbable" climb as weve already undertaken for the vitreous mass eye of the worm. Lenses, like eyes themselves, seem to have evolved many times independently. "Mount Improbable" has many peaks and hillocks.
Retinas, too, betray their manifold origins by their variable forms. With one exception, all the eyes I have so far illustrated have had their photocells in front of the nerves connecting them to the brain. This is the obvious way to do it, but it is not universal. The flatworm in Figure 5.4a keeps its photocells apparently on the wrong side of their connecting nerves. So does our own vertebrate eye. The photocells point backwards, away from the light. This is not as silly as it sounds. Since they are very tiny and transparent, it doesnt much matter which way they point: most photons will go straight through and then run the gauntlet of pigment-laden baffles waiting to catch them. The only sense in which it even means much to say that vertebrate photocells point backwards is that the wires (nerves) connecting them to the brain depart in the wrong direction, towards the light rather than towards the brain. They then run over the front surface of the retina towards one particular place, the so-called blind spot. This is where they dive through the retina into the optic nerve, which is why the retina is blind at this spot. Although we are all technically blind at the spot, we scarcely know it because the brain is so clever at reconstituting the missing bit. We only notice the blind spot if the image of some small discrete object, which we have independent evidence exists, moves on to it: it then appears to go out like a light, apparently replaced by the general background color of the area.
Ive said that it makes little difference if the retina is back-to-front. A case could be made that, absolutely all other things being equal, it might have been better if our retinas were the right Way round. It is perhaps a good example of the fact that "Mount Improbable" has more than one peak, with deep valleys between. Once a good eye has started to evolve with its retina back-to-front, the only way to ascend is to improve the present design of eye. Changing to a radically different design involves going downhill, not just a little way but down a deep chasm, and that is not allowed by natural selection. The vertebrate retina faces the way it does because of the way it develops in the embryo, and this certainly goes back to its ancient ancestors. The eyes of many invertebrates develop in different ways, and their retinas are consequently the right way round.
Setting aside the interesting fact of their pointing backwards, vertebrate retinas scale some of the loftiest peaks on the mountain. The human retina has about 166 million photocells, divided into various kinds. The basic division is into rods (specialized for low-precision, non color vision at relatively low light levels) and cones (specialized for high-precision color vision in bright light). As you read these words you are using only cones. If Juliet had seen Halleys Comet, it would have been her rods that were responsible. The cones are concentrated in a small central area, the fovea (you are reading with your foveas) where there are no rods. This is why, if you want to see a really dim object like Halleys Comet, you must point your eyes not directly at it but slightly away, so that its meagre light is off the fovea. Numbers of photocells, and differentiation of photocells into more than one type, present no special problems from the point of view of climbing "Mount Improbable". Both kinds of improvement obviously constitute smooth gradients up the mountain.
Big retinas see better than small retinas. You can fit more photocells in, and you can see more detail. But, as always, there are costs. Remember the surrealist snail of Figure 5.1. But there is a way in which a small animal can, in effect, enjoy a larger retina than it pays for. Professor Michael Land of Sussex University, who has an enviable track record for exotic discoveries in the world of eyes and from whom I have learned much of what I know about eyes, found a wonderful example in jumping spiders. No spiders have compound eyes: jumping spiders have taken the camera eye up to a remarkable peak of economy (Figure 5.17). These engaging little animals, whose habit of cocking their heads to look at you gives them an almost human charm, stalk their prey like a cat and then jump on to i� explosively and without warning. Explosive it more or less literally is, by the way, for they jump by hydraulically pumping fluid into all eight legs simultaneously - a little like the way we (those of us who have them) erect our penises, but their leg erections are sudden rather than gradual.
What Land discovered was an extraordinary retina. Instead of being a wide sheet on which a full image can be projected, it is a long, vertical strip , not wide enough to accommodate a decent image. But the spider makes up for the narrowness of its retina by an ingenious makeshift. It moves its retina systematically about, scanning the area where an image might be projected. Its effective retina is, therefore, much larger than its actual retina - rather on the same principle as the bolas spider with its whirling single thread approximates the catchment area of a proper web. If the jumping spiders retina finds an interesting object, such as a moving fly or another jumping spider, it concentrates its scanning movements in the precise area of the target. This gives it the dynamic equivalent of a fovea. Using this clever trick, jumping spiders have carried the lens eye to a respectable little peak in their local area of "Mount Improbable".
I introduced the lens as an excellent remedy for the shortcomings of the pinhole eye. It isnt the only one. A curved mirror constitutes a different principle from a lens, but it is a good alternative solution to the same problem of gathering a large amount of light from each point on an object, and focusing it to a single point on an image. For some purposes a curved mirror is actually a more economical solution to the problem than a lens, and the biggest optical telescopes in the world are all reflectors (Figure 5.18a). A minor problem with a reflecting telescope is that the image is formed in front of the mirror, actually in the pathway of the incoming rays. Reflecting telescopes usually have a small mirror to reflect the focused image sideways into an eyepiece or a camera.
The small mirror doesnt get in the way, right enough to spoil the image, anyway. No focused image of the little mirror is seen: it merely causes a small reduction in the total amount of light hitting the big mirror at the back of the telescope. The curved mirror, then, is a theoretically workable physical solution to an important problem. Are there any examples of curved mirror eyes ,in the animal kingdom? The earliest suggestion along these lines was made by my old Oxford Professor, Sir Alister Hardy, commenting on his painting of a remarkable deep-sea crustacean called Gigantocypris (Figure 5.18b). Astronomers capture what few photons arrive from distant stars with huge curved mirrors in observatories like Mount Wilson and Palomar. It is tempting to think that Gigantocypris is doing the same thing with the few photons that penetrate the deep oceans, but recent investigations by Michael Land rule out any resemblance in detail. it is at the moment not clear how Gigantocypris sees.
There is another kind of animal, however, that definitely uses a bona fide curved mirror to form an image, albeit it has a lens to help. Once again, it was discovered by that King Midas of animal eye research, Michael Land. The animal is the scallop. The photograph in Figure 5.18c is an enlargement of.a small piece (two shell-corrugations in width) of the gap of one of these bivalves. Between the shell and the tentacles is a row of dozens of little eyes. Each eye forms an image, using a curved mirror which lies well behind the retina. It is this mirror that causes each eye to glow like a tiny blue or green pearl. In section, the eye looks like Figure 5.18d. As I mentioned, there is a lens as well as a mirror, and Ill come back to this. The retina is the whole grayish area lying between the lens and the curved mirror. The part of the retina which sees the sharp image projected by the mirror is the portion tightly abutting the back of the lens. That image is upside-down and it is formed by rays reflected backwards by the mirror.
So, why is there a lens at all? Spherical mirrors like this one are subject to a particular kind of distortion called spherical aberration. A famous design of reflecting telescope, the Schmidt, overcomes the problem by a cunning combination of lens and mirror. Scallop eyes seem to solve the problem in a slightly different way. Spherical aberration can theoretically be overcome by a special kind of lens whose shape is called a Cartesian oval. Figure 5.18e is a diagram of a theoretically ideal Cartesian oval. Now look again at. the profile of the actual lens of the scallop eye (Figure 5.18d). On the basis of the striking resemblance, Professor Land suggests that the lens is there as a corrector for the spherical aberration of the mirror which is the main image-forming device.
As for the origin of the curved mirror eye on the lower slopes of its region of the mountain, we can make an educated guess. Reflecting layers behind retinas are common in the animal kingdom, but for a different purpose, not image forming, as in scallops. If you go out into the ", woods with a bright spotlight you will see numerous twin beams glaring ,, straight back at you. Many mammals, especially nocturnal ones like golden potto or angwantibo from West Africa , have a tapetum, a reflecting layer behind the retina. What the tapetum does is provide a second chance of catching photons that the photocells failed to stop: each photon is reflected straight back to the very photocell that missed it coming the other way, so the image is not distorted. Invertebrates, they too, have discovered the tapetum. A bright torch in the woods is an excellent way to find certain kinds of spider. Indeed, looking at the portrait of a wolf spider (Figure 5.19a), you wonder why the cats eyes that mark our roads are not called spider eyes. Tapetums for capturing every last photon may well have evolved in ancestral cup eyes before lenses. Perhaps the tapetum is the pre-adaptation which, in a few isolated creatures, has become modified to form a reflecting telescope kind of eye. Or the mirror may have arisen from another source. It is hard to be sure. Lenses and curved mirrors are two ways of sharply focusing an image. In both cases the image is upside-down and left-right reversed. A completely different kind of eye, which produces an image the right way up, is the compound eye, favored by insects, crustaceans, some worms and mollusks, king crabs (strange marine creatures said to be closer to spiders than to real crabs) and the large group of now extinct trilobites. Actually there are several different kinds of compound eye. Ill begin with the most elementary kind, the so-called apposition compound eye. To understand how the apposition eye works, we go back nearly to the bottom of "Mount Improbable". As we have seen, if you want an eye to see an image or indeed go beyond signalling the mere intensity of light, you need more than one photocell, and they must pick up light from different directions. One way to make them look in different directions is to place it far from them in a cup, backed by an opaque screen. All the eyes we have sofar talked about have been descendants of this concave cup principle. But perhaps an even more obvious solution to the problem is to place the photocells on the convex outside surface of a cup, thereby causing them to look outwards in different directions. This is a good way to think of a compound eye, at its simplest.
Remember when we first introduced the problem of forming an image of a dolphin. I pointed out that the problem could be regarded as the problem of having too many images. An infinite number of dolphins on the retina, every way up and in every position on the retina, adds up to no visible dolphin at all (Figure 5.20a). The pinhole eye worked because it filtered out almost all the rays, leaving only th~ minority that cross each other in the pinhole and form a single upside-down image of the dolphin. We treated the lens as a more sophisticated version of the same principle. The apposition compound eye solves the problem in an even simpler way.
The eye is built as a dense cluster of long straight tubes, radiating out in all directions from the roof of a dome. Each tube is like a gun sight which sees only the small part of the world in its own direct line of fire. In terms of our filtering metaphor, we could say that rays coming from other parts of the world are prevented, by the walls of the tube and the backing of the dome, from hitting the back of the tube where the photocells are.
Thats basically how the apposition compound eye works. In practice, each of the little tube eyes, called an ommatidium (plural ommatidia), is a bit more than a tube. It has its own private lens, and its own tiny retina of, usually, half a dozen or so photocells. Insofar as each ommatidium produces an image at all at the bottom of the narrow tube, that image is upside-down: the ommatidium works like a long, poor-quality, camera eye. But the upside-down images of the individual ommatidia are ignored. The ommatidium reports only how much light comes down its tube. The lens serves only to gather more light rays from the ommatidiums gun sight direction and focus them on to the retina. When all the ommatidia are taken together, their summed image is the right way up, as shown in Figure 5.20b.
As always, image doesnt have to mean what we humans would think of as an image: an accurate, Technicolor perception of an entire scene. Instead, we are talking about any kind of ability to use the eyes to distinguish what is going on in different directions. Some insects might, for example, use their compound eyes only to track moving targets. They might be blind to still scenes. The question of whether animals see things in the same way as we do is partly a philosophical one and it may be a more than usually difficult task trying to answer it.
The compound-eye principle works well enough for, say, a dragonfly zeroing in on a moving fly but, in order for a compound eye to see as much detail as we see, it would need to be hugely bigger than the kind of simple camera eye that we possess. Here is approximately why this is. Obviously, the more ommatidia you have, all looking in slightly different directions, the more fine detail you can see. A dragonfly may have 30,000 ommatidia and it is pretty good at hawking insects on the wing (Figure 5.21). But in order to see as much detail as we can see, youd need millions of ommatidia. The only way to fit in millions of ommatidia is to make them exceedingly tiny. And unfortunately there is a strict limit on how small an ommatidium can be. It is the same limit as we met in talking about very small pinholes, and it is called the diffraction limit. The consequence is that, in order to make a compound eye see as precisely as the human camera eye, .the compound eye would have to be ludicrously large: twenty-four meters in diameter! The German scientist Kuno Kirschfeld dramatized this by drawing what a man would look like if he could see as well as a normal man can see, but using compound eyes (Figure 5.22). The honeycomb pattern on the drawing is impressionistic, by the way. Each facet drawn actually stands for 10000 ommatidia. The reason the mans compound eyes are only about one meter across instead of twenty-four is that Kirschfeld made allowance for the fact that we humans see very precisely only in the center of our retina. He took an average of our precise central vision and our much less precise vision towards the edges of our retina, and came up with the one meter eye shown. Whether one ere across or twenty-four, a compound eye this large is impractical.
The moral is, if you want to see precise detailed images of the world, use a simple camera eye with a single, good lens, not a compound eye. Dan Nilsson even remarks of compound eyes that "It is only a small exaggeration to say that evolution seems to be fighting a desperate battle to improve a basically disastrous design."
Why, then, dont insects and crustaceans abandon the compound eye and evolve camera eyes instead? It may be one of those cases of becoming trapped the wrong side of a valley on the massif of "Mount Improbable". To change a compound eye into a camera eye, there has to be a continuous series of workable intermediates: you cannot travel down into a valley as a prelude to mounting a higher peak. So what is the problem about intermediates between a compound eye and a camera eye?
At least one outstanding difficulty comes to mind. A camera eye forms an upside-down image. A compound eyes image is the right way up. Finding an intermediate between those two is a tough proposition, to put it mildly. A possible intermediate is no image at all. There are some animals, living in the deep sea or otherwise in near total darkness, who have so few photons to play with that they give up on images altogether. All that they can hope for is to know whether there is light at all. An animal such as this could lose its image-processing nervous apparatus altogether and hence be in a position to make a fresh start up a completely different slope of the mountain. It could therefore constitute an intermediate on the path from a compound eye to a camera eye.
Some deep-sea crustaceans have large compound eyes but no lenses or optical apparatus at all. Their ommatidia have lost their tubes and their photocells are exposed right at the outer surface of where they will pick up what few photons there are, regardless of direction. From there it would seem but a small step to the remarkable eye of Figure 5.23. It belongs to a crustacean, called Ampelisca, which doesnt live particularly deep - perhaps it is on the way back up again from deep-sea ancestors. Ampeliscas eye works as a camera eye, with a single lens forming an upside-down image on a retina. But the retina is clearly derived from a compound eye and consists of the remains of a bank of ommatidia. A small step, maybe, but only if, during the interregnum of near total blindness, the brain had enough evolutionary time to forget all about processing right-way-up images.
That is an example of evolution from compound eye to camera eye (yet another example, by the way, of the case with which eyes seem to evolve independently all around the animal kingdom). But how did the compound eye evolve in the first place? What do we find on the lower slopes of this particular peak of "Mount Improbable"?
Once again we may be helped by looking around the modem animal kingdom. Outside the arthropods (insects, crustaceans and their kin), compound eyes are found only in some Polychaet worms (ragworms and tubeworms) and in some bivalve mollusks (again, presumably independently evolved). The worms and mollusks are helpful to us as evolutionary historians because they also include among their number some primitive eyes which look like plausible intermediates strung out along the lower slopes of "Mount Improbable" leading to a compound-eye peak. The eyes in Figure 5.24 come from two different worm species. Once again, these are not ancestors, they are modern species and they are probably not even descended from the true intermediates. But they could easily be giving us a glimpse of what the evolutionary progression might have been like, from a loose clustering of photocells on the left to a proper compound eye on the right. This slope is surely just as gentle the one we strolled up to reach the ordinary camera eye.
Ommatidia, as we have so far discussed them, depend for their effectiveness on being isolated from their neighbours. The gunsight that is looking at the dolphins tailtip must not pick up rays from other parts of the dolphin, or we should be back with our original problem of millions of dolphin images. Most ommatidia achieve isolation by having asheath of dark pigment around the tube. But there are times when this has bad side effects. Some sea creatures rely on transparency for their camouflage. They live in sea water and they look like sea water. The essence of their camouflage, then, is to avoid stopping photons. Yet the whole point of dark screens around ommatidia is to stop photons. How to escape from this cruel contradiction?
There are some deep-sea crustaceans who have come up with an ingenious partial solution (Figure 5.25). They dont have screening pigment, and their ommatidia are not tubes in the ordinary sense. Instead they are transparent light guides, working just like man-made fibrous optic systems. Each light guide swells, at its front end, into a tiny lens, of varying refractive index like a fish eye. Lens and all, the light guide as a whole concentrates a large amount of light on to the photocells at base. But this includes only light coming from straight in the line of gun sight. Beams coming sideways at a tube, instead of being shielded by, a pigment screen, are reflected back and dont enter the light guide.
Not all compound eyes even try to isolate their private supply of ". light. It is only eyes of the apposition type that do. There are at least three different kinds of superposition compound eye which do something more subtle. Far from trapping rays in tubes or fibre-optic light guides, they allow rays that pass through the lens of one ommatidium to be picked up by a neighboring ommatidiums photocells. There is an empty, transparent zone, shared by all ommatidia. The lenses of all ommatidia conspire together to form a single image on a shared retina which is jointly put together from the light-sensitive cells of all the ommatidia. Inset Figure 5.26 is Michael Lands picture of Charles Darwin, seen through the compound lens of a fireflys superposition compound eye.
The image in a superposition compound eye, like that of apposition compound eyes but unlike that of camera eyes or that of Figure 5.23 Ampelisca, is the right way up. This is what youd expect, assuming that superposition eyes are derived from apposition ancestors. It makes historical sense, and it must have made for an effortless transition as far as the brain was concerned. But it is still a very remarkable fact. For consider the physical problems of constructing a single right-way-up image in this way. Each individual ommatidium in an apposition eye has a normal lens in front of it which, if it makes an image at all, makes an upside-down one. To convert an apposition eye into a superposition one, therefore, the rays, as they pass through each lens, have somehow to be turned the right way up. Not only this, all the separate images from the different lenses have to be carefully superimposed to give one shared image. The advantage of doing this is that the shared image is much brighter. But the physical difficulties of turning the rays round are formidable. Amazingly, not only has the problem been solved in evolution, it has been solved in at least three independent ways: using fancy lenses, using fancy mirrors and using fancy neural circuitry. The details are so intricate that to spell them out would imbalance this already quite complicated chapter, and Ill deal with them only briefly.
A single lens turns the image upside-down. By the same token another lens, a suitable distance behind it, would turn it the right way up again. The combination is used in an instrument called a Keplerian telescope. The equivalent effect can be achieved in a single complex lens, using fancy gradations of refractive index. As we have seen, living lenses, unlike man-made ones, are good at achieving gradations of refractive index. This method of simulating the effect of a Keplerian telescope is used by mayflies, lacewings, beetles, moths, caddises and members of five different groups of crustaceans. The distance of their cousin ship suggests that at least several of these groups evolved the same Keplerian trick independently of one another. An equivalent trick is pulled by three groups of crustaceans, using mirrors. Two of these three groups also contain members that do the lens trick. Indeed, if you look at which animal groups have adopted which of the several different kinds of compound eye, you notice a fascinating thing. The different solutions to problems pop up here, there and everywhere, suggesting, yet again, that they evolve rapidly and at the drop of a hat.
Neural superposition or wired-up superposition has evolved in the large and important group of two-winged insects, the flies (fireflies, by the way, such as those mentioned in Figure 5.26, are not flies at all, but beetles). A similar system occurs in water boatmen, where it seems to have evolved - yet again - independently. Neural superposition is fiendishly cunning. In a way it shouldnt be called superposition at all, because the ommatidia are isolated tubes just as in apposition eyes. But they achieve a superposition-like effect by ingenious wiring of nerve cells behind the ommatidia. Heres how. Youll remember that the retina of a single ommatidium is made up of about half a dozen photocells. In ordinary apposition eyes, the firing of all six photocells is simply added together, which is why I put retina in quotation marks: all photons that shoot down the tube are counted, regardless of which photocell they hit. The only point of having several photocells is to increase the total sensitivity to light. This is why it doesnt matter that the tiny image at the bottom of an apposition ommatidium is technically upside-down.
But in the eye of a fly the outputs of the six cells are not pooled with each other. Instead, each one is pooled with the outputs of particular cells from neighboring ommatidia (Figure 5.27). In the interests of clarity, the scale is all wrong in this diagram. For the same reason, the arrows dont represent rays (which would be bent by the lenses) but mappings from points on the dolphin to points in the bottoms of tubes. Now, see the shattering ingenuity of this scheme. The essential idea is that those photocells that are looking at the head of the dolphin in one ommatidium are ganged together with those photocells that are looking at the head of the dolphin in neighboring ommatidia. Those photocells that are looking at the tail of the dolphin in one ommatidium are wired up together with those photocells that are looking at the tail of the dolphin in neighboring ommatidia. And so on. The result is that each bit of the dolphin is being signalled by a larger number of photons than there would be in an ordinary apposition eye with a simple tube arrangement. It is a kind of computational, rather than an optical, solution to our old problem of how to augment the number of photons arriving from any one point on our dolphin.
Nilssons estimate for the rate of evolution of a camera eye was, you will recall, that it was by geological standards more or less instantaneous. Youd be lucky to find fossils that recorded the transitional stages. Exact estimates have not been done for compound eyes or any of the other designs of eye, but I doubt if theyd be significantly slower. One doesnt ordinarily expect to be able to see the details of eyes in fossils, because they are too soft to fossilize. Compound eyes are an exception because much of their detail is betrayed in the elegant array of more or less horny facets on the outer surface. Inset Figure 5.28 shows a trilobite eye, from the Devonian era, nearly 400 million years ago. It looks just as advanced as a modem compound eye. This is what we should expect if the time it takes to evolve an eye is negligible by geological standards.
A central message of this chapter is that eyes evolve easily and fast, at the drop of a hat. I began by quoting the conclusion of one authority that eyes have evolved independently at least forty times in different parts of the animal kingdom. On the face of it, this message might seem challenged by an intriguing set of experimental results, recently reported by a group of workers in Switzerland associated with Professor Walter Gehring. I shall briefly explain what they found, and why it does not really challenge the conclusion of this chapter. Before I begin, I need to apologize for a maddeningly silly convention adopted by geneticists over the naming of genes. The gene called eyeless in the fruit fly Drosophila actually makes eyes! (Wonderful, isnt it?) The reason for this wantonly confusing piece of terminological contrariness is actually quite simple, and even rather interesting. We recognize what a gene does by noticing what happens when it goes wrong. There is a gene which, when it goes wrong (mutates), causes flies to have no eyes. The position on the chromosome of this gene is therefore named the eyeless locus (locus is the Latin for place and it is used by geneticists to mean a slot on a chromosome where alternative forms of a gene sit). But usually when we speak of the locus named eyeless we are actually talking about the normal, undamaged form of the gene at that locus. Hence the paradox that the eyeless gene makes eyes. It is like calling a loudspeaker a silence device because you have discovered that, when you take the loudspeaker out of a radio, the radio is silent. I shall have none of it. I am tempted to rename the gene eye maker, but this would be confusing too. I shall certainly not call it eyeless and shall adopt the recognized abbreviation eye.
Now, it is a general fact that although all of an animals genes are present in all its cells, only a minority of those genes are actually turned on or expressed in any given part of the body. This is why livers are different from kidneys, even though both contain the same complete set of genes. In the adult Drosophila, eye usually expresses itself only in the head, which is why the, eyes develop there. George Halder, Patrick Callaerts and Walter Gehring discovered an experimental manipulation that led to eyes being expressed in other parts of the body. By doctoring Drosophila larvae in cunning ways, they succeeded in making eye express itself in the antennae, the wings and the legs. Amazingly, the treated adult flies grew up with fully formed compound eyes on their wings, legs, antennae and elsewhere (Inset Figure 5.29). Though slightly smaller than ordinary eyes, these ectopic eyes are proper compound eyes with plenty of properly formed ommatidia. They even work. At least, we dont know that the flies actually see anything through them, but electrical recording from the nerves at the base of the ommatidia shows that they are sensitive to light.
That is remarkable fact number one. Fact number two is even m remarkable. There is a gene in mice called small eye and one in hum called aniridia. These, too, are named using the geneticists negative vention: mutational damage to these genes causes reduction or ab of eyes or parts of eyes. Rebecca Quiring and Uwc Waldorf, working, the same Swiss laboratory, found that these particular mammal genes were almost identical, in their DNA sequences, to the eye gene in Drosophilias. This means that the same gene has come down from remote ancestors modem animals as distant from each other as mammals and insects. Moreover, in both these major branches of the animal kingdom the gene seems to have a lot to do with eyes. Remarkable fact number three is almost too startling. Halder, Callaerts and Gehring succeeded in introducing the mouse gene into Drosophila embryos. Mirabile dictu, the mouse gene induced ectopic eyes in Drosophila. Inset Figure 5.29 (bottom), shows a small compound eye induced on the leg of a fruitfly by the mouse equivalent of ey. Notice, by the way, that it is an insect compound eye that has been induced, not a mouse eye. The mouse gene has simply switched on the eyemaking developmental machinery of Drosophila. Genes with pretty much the same DNA sequence as ey have been found also in molluscs, marine worms called nemertines, and sea-squirts. Ey may very well be universal among animals, and it may rum out to be a general rule that a version of the gene taken from a donor in one part of the aminal kingdom can induce eyes to develop in recipients in an exceedingly remote part of the animal kingdom.
What does this spectacular series of experiments mean for our conclusion in this chapter? Were we wrong to think that eyes have developed forty times independently? I dont think so. At least the spirit of the statement that eyes evolve easily and at the drop of a hat remains unscathed. These experiments probably do mean that the common ancestor of Drosophila, mice, humans, sea-squirts and so on had eyes. The remote common ancestor had vision of some kind, and its eyes, whatever form they may have taken, probably developed under the influence of a sequence of DNA similar to modern ey. But the actual form of the different kinds of eye, the details of retinas and lenses or mirrors, the choice of compound versus simple, and if compound the choice among apposition or various kinds of superposition, all these evolve independently and rapidly. We know this by looking at the sporadic - almost capricious - distribution of these various devices and systems, dotted around the animal kingdom. In brief, animals often have an eye that resembles their remoter cousins more than it resembles their closer cousins, The conclusion remains unshaken by the demonstration that the common ancestor of all these animals probably had eyes of some kind, and that the embryonic development of all eyes seems to have enough in corn mon to be inducible by the same DNA sequence.
After Michael Land had kindly read and criticized the first draft of this chapter, I invited him to attempt a visual representation of the eye region of "Mount Improbable" and Figure 5.30 shows what he drew. It is in the nature of metaphors that they are good for some purposes but not others and we must be prepared to modify them, or even drop them altogether, when necessary. This is not the first occasion when the reader will have noticed that "Mount Improbable", for all that it has a singular name like theJungfrau, is actually a more complicated, multiple-peaked affair.
That other great authority on animal eyes, Dan Nilsson, who also read the chapter in draft, summed up the central message by calling my attention to what may be the most bizarre example of the ad hoc and opportunistic evolution of an eye. Three times independently, in three different groups of fish, the so-called four-eyed condition has evolved. Probably the most remarkable of the four-eyed fish is Bathylychnops exilis (Figure 5.31). It has a typical fish eye looking outwards in the usual direction. But a secondary eye has evolved in addition, lodged in the wall of the main eye and looking straight downwards. What it looks at, who knows? Perhaps Bathylychnops suffers from a terrible predator with the habit of approaching from below. From our point of view the interesting thing is this. The embryological development of the secondary eye is completely different from that of the main eye, although we may surmise that its development may turn out to be induced in nature by a version of the ey gene. In particular, as Dr Nilsson put it in his letter to me, This species has re-invented the lens despite the fact it already had one. It serves as a good support for the view that lenses are not difficult to evolve. Nothing is as difficult to evolve as we humans imagine it to be.
Darwin gave too much when he bent over backwards to concede the difficulty of evolving an eye. And his wife took too much when she underlined her scepticism in the margin. Darwin knew what he was doing. Creationists love the quotation that I gave at the beginning of this chapter, but they never complete it. After making his rhetorical concession, Darwin went on: "When it was first said that the sun stood still and the world turned round, the common sense of mankind declared the doctrine false; but the old saying of Vox populi, vox Dei, as every philosopher knows, cannot be trusted in science. Reason tells me, that if numerous gradations from an imperfect and simple eye to one perfect and complex, each grade being useful to its possessor, can be shown to exist, as is certainly the case; if further, the eye ever slightly varies, and the variations be inherited, as is likewise certainly the case; and if such variations should ever be useful to any animal under changing conditions of life, then the difficulty of believing that a perfect and complex eye could be formed by natural selection, though insuperable by our imagination, cannot be considered real.
Created on ... April 17, 2004
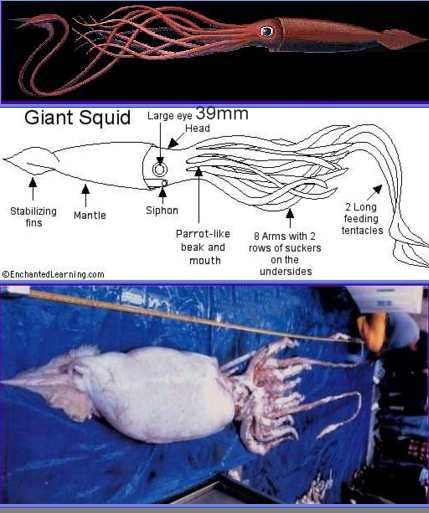{kind=link}