|
|
| |
I consider the results of the statistic study, included in MeMint, with the longitudinal data of the Young Adulthood Study, 1939-1967 to be conclusive about the genetic influence on intelligence (r2 till 0,99), the significance of the less powerful gene, important functionalities of the sexual diversification and about the existence of a teleological evolution. |
|
| |
|
|
||||||
|
|
|
Translate Last update: September 2002 |
GENERAL THEORY OF THE CONDITIONAL EVOLUTION OF LIFE |
||
|
||||||
|
||||||
|
|
|||||
|
||||||
|
|
|||||
|
||||||
|
|
|||||
|
||||||
|
|
|||||
|
||||||
|
|
|||||
| ||||||
|
|
|||||
|
||||||
|
|
|||||
|
||||||
|
|
|||||
|
This is a translation of a reduce version of the GTCEL e-book. (The full version is in Spanish) |
|
|
|

|
|
|
NOTE: The model of the inheritance of intelligence has been validated using the longitudinal data set of the Young Adulthood Study, 1939-1967.
|
|
|
|
|
The proposed model assumes the following hypotheses:
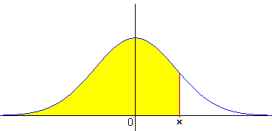
In order to facilitate the understanding of the model and its statistical analysis, we are going to choose the controverted and very studied subject of the inheritance of intelligence. Numerous studies based on measurements of the individual IQ or coefficients of intelligence exist. The relation to other individuals estimates the generally accepted IQs, although many authors doubt these measurements and the unique concept of intelligence. So, the IQ refer to the relative position defined by means of a standardised function x (I) of the statistical distribution of the IQ studied for the process of validation of this function. The studies have some contradictory conclusions, whereas in studies with identical twins a correlation of 80-85% is reached, for other types of familiar relations, decreases to a 30%. For me, the conclusion is that intelligence it is inherited as it demonstrates the high correlation between identical twins. The low correlation in the rest of the cases is due to the incorrect definition of the form in which the inheritance is transmitted in agreement with the exposition of the GTCEL. This figure shows the shape of the Normal function x(IQ), which we are going to use. For each IQ value, the function indicates the acumulated probability that the IQ of the population is equal or less than the first one. For example x(100) = 0.5 and the opposite function xinv(Prob) = IQ, that means, xinv(0.5) = 100. |
|
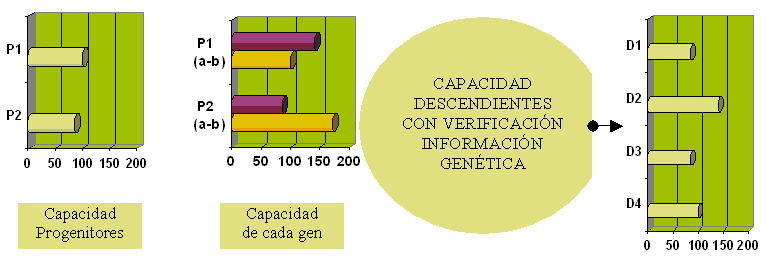
According with the values of the previous figure and using the Catell scale, mean 100 and standard deviation 24, we will have the following percentils: |
| Progenitor o Descendent | Significant gen with VGI | Percentil = x (IQ) |
| P1 | P1b = 100 | 0.50 |
| P2 | P2a = 85 | 0.27 |
| D1 | P2a = 85 | 0.27 |
| D2 | P1a = (*) | 0.95 |
| D3 | P2a = 85 | 0.27 |
| D4 | P1b = 100 | 0.50 |
|
|
|
|